
di Mario Pagano
Introduzione
Le foglie sono organi essenziali per mantenere un corretto equilibrio tra O2 e CO2 in atmosfera attraverso i processi di fotosintesi e respirazione (Bhardwaj et al., 2013). A tal riguardo, un aspetto fisiologico, condiviso anche da altri organismi viventi, è l’alta ramificazione e la diversità strutturale del sistema vascolare (Parsons-Wingerter et al., 2011; Sack et al., 2012).
La regolazione nell’efficienza del trasporto dell’acqua (cioè la massa d’acqua trasportata per unità di gradiente di pressione) è evidente in tutto il percorso evolutivo della foglia. La struttura dello xilema, in grado di aumentare l’efficienza idraulica e di mantenere la continuità della colonna d’acqua, è cruciale per la sopravvivenza delle specie vegetali. In particolare, il sistema vascolare della foglia ha due funzioni principali: il trasporto di soluzioni acquose e il supporto meccanico. L’acqua, soluti e gli ormoni sono trasportati nello xilema mentre i carboidrati sono trasportati attraverso il floema (Roth-Nebelisck et al., 2001).
L’architettura della vena, oltre ad essere essenziale per un supporto meccanico della foglia, può anche giocare un ruolo cruciale nell’efficienza della fotosintesi (Brodribb et al., 2007). Inoltre, studi recenti, hanno dimostrato come l’intensità cromatica del verde della pagina fogliare, descritta da un indice di colore afferente allo spettro RGB (red-green-blu), sia influenzata da numerosi fattori come ad esempio il bilancio di carbonio nell’ecosistema e l’efficienza della fotosintesi (Ahrends et al., 2009; Richardson et al., 2007). A tale riguardo, l’influenza dell’ambiente sullo spettro RGB, può essere eliminata utilizzando i seguenti indici (Sonnentag et al., 2012):

Durante gli ultimi anni, come ampiamente documentato dalla letteratura scientifica, sono stati sviluppati numerosi indici per le immagini digitali (Meyer et. al., 2008; Perez et. al., 2000; Woebbecke et al., 1995). L’obiettivo di molti di questi studi è stato quello di mettere appunto tecniche idonee sia per separare l’immagine delle piante dal rispettivo sfondo (Meyer et al., 2008; Perez et al., 2000; Woebbecke et al., 1995; Sonnentag et al., 2012), così da garantirne una identificazione automatica delle stesse, che quello di evidenziare con accuratezza lo spettro RGB.
Nella presente ricerca, invece, sono state utilizzate tecniche di analisi dell’immagine con la finalità di porre in evidenza una correlazione esistente tra il sistema vascolare delle foglie e la temperatura della pagina fogliare.
Materiali e metodi
La ricerca ha interessato la specie Vitis vinifera (L.) (cultivar ‘Gorgottesco’). Tutte le viti utilizzate nella ricerca sono state esposte alle stesse condizioni agronomiche ed ambientali (27,1 °C; 64,1 % di umidità relativa) all’interno della serra del CREA di Arezzo, Italia.
A tal proposito, sono state individuate le foglie con un età corrispondente ad un periodo con un’efficienza massima della fotosintesi (Kriedemann et al., 1970). A tal riguardo, sono stati eseguiti sulle foglie, ancora posizionate sulle piante, i rilievi della temperatura utilizzando un termometro ad infrarossi (Fluke 62 Mini). Il termometro è stato posizionato verticalmente sul campione da analizzare. Le foglie sono state campionate tra il sesto e il settimo nodo di ogni tralcio principale. Successivamente, sono state acquisite direttamente sulla pagina fogliare, utilizzando una fotocamera EOS 700D (Canon, Giappone) equipaggiata con un obiettivo macro EF 100 millimetri f/2,8 (Canon, Giappone) ed un microscopio digitale Dinolite (AM-313T5, Anmo Electronics Corporation, Hsinchu, Taiwan), le immagini del sistema vascolare.
L’acquisizione digitale del campione è stata eseguita dividendo l’area della foglia in quadranti di 1 mm2. In ogni quadrante sono state incluse le vene dal 2° ordine in poi. Le immagini sono state elaborate attraverso l’uso del software ImageJ 1.49v (National Institutes of Health, USA), mentre con l’ausilio del microscopio Dinolite sono stati effettuati degli ingrandimenti delle porzioni di lamina ritenute più interessanti per la realizzazione di modelli 3D. Il modello 3D è stato realizzato utilizzando il software ImageJ. Questo modello è una possibilità offerta da questo software per simulare la distribuzione del calore sul sistema vascolare della foglia.
Risultati e discussione
La tecnica di elaborazione delle immagini utilizzata in questo lavoro ha permesso di ottenere informazioni sul sistema vascolare della vite. Dall’analisi statistica dei dati raccolti in ogni singolo quadrante della pagina fogliare, è emersa una correlazione significativa tra a densità delle vene e la temperatura della lamina (fig. 1). Questa correlazione supporta l’ipotesi che il sistema vascolare delle foglie, con particolare riferimento a quelle di vite, può rivestire un ruolo nel processo di raffreddamento della medesima. Infatti, ad una maggiore densità delle vene corrisponde una temperatura inferiore.
Inoltre, è stata trovata una correlazione significativa tra l’indice di verde (gcc) e la temperatura della pagina fogliare (fig. 2). La correlazione evidenzia un più alto valore dell’indice gcc in funzione di una temperatura inferiore. Probabilmente, questa informazione può essere associata ad un buon stato di salute delle foglie. Inoltre, è emersa anche una significativa correlazione tra la densità delle vene e l’indice gcc (fig. 3). Un indice gcc maggiore può, probabilmente, essere abbinato ad un più alto contenuto di clorofilla. Infine è stato costruito un modello virtuale 3D LUT (fig. 4) per evidenziare il profilo termico del sistema vascolare.
Conclusioni
Lo studio dell’architettura della vena è il punto principale per comprendere le dinamiche dei fluidi coinvolti nei tessuti vascolari. Il presente lavoro è un contributo per evidenziare la correlazione esistente tra il sistema vascolare della foglia e la temperatura.
I dati preliminari evidenziano una correlazione tra la densità del sistema vascolare e la temperatura della lamina. I risultati confermano l’ipotesi che la rete venosa è correlata con la temperatura della medesima. Questa correlazione può assumere un ruolo nel raffreddamento della foglia. I passi successivi saranno orientati nel confermare queste informazioni preliminari utilizzando una telecamera termica. In futuro, sarà possibile, quindi, realizzare modelli predittivi in grado di integrare le conoscenze acquisite attraverso la “lettura delle foglie” come, ad esempio, evidenziare il potenziale rischio di stress idrico e di conseguenza le esigenze di irrigazione del vigneto.
Bibliografia
- Ahrends H.E., Etzold S., Kutsch W.L., Stoeckli R., Bruegger R., Jeanneret F., Wanner H., Buchmann N., Eugster W. 2009. Tree phenology and carbon dioxide fluxes: use of digital photography at for process-based interpretation the ecosystem scale. Climate Research, 39: 261–274.
- Bhardwaj A., Kaur M., Kumar A. 2013. Recognition of plants by leaf image using moment invariant and texture analysis. International Journal of Innovation and Applied Studies, 1: 237-248.
- Brodribb T.J., Field T.S., Jordan G.J. 2007. Leaf Maximum Photosynthetic Rate and Venation Are Linked by Hydraulics. Plant Physiology, 144: 1890-1898.
- Kriedemann P.E., Kliewer W.M., Harris J.M. 1970. Leaf age and photosynthesis in Vitis vinifera (L.). Vitis 9: 97-104.
- Meyer, G.E., Neto, J.C., 2008. Verification of color vegetation indices for automated crop imaging applications. Computers and Electronics in Agriculture, 63: 282–293.
- Pagano M., Storchi P. 2015. Leaf vein density: a possible role as cooling system. Journal of Life Sciences (ISSN 1934-7391). In stampa.
- Parsons-Wingerter P., Vickerman M.B. 2011. Informative Mapping By VESGEN Analysis of Venation Branching Pattern in Plant Leaves Such as Arabidopsis thaliana. Gravitational and Space Biology, 25: 69-71.
- Perez A.J., Lopez F., Benlloch J.V., Christensen S. 2000. Colour and shape analysis techniques for weed detection in cereal fields. Computers and Electronics in Agriculture, 25: 197–212.
- Richardson, A.D., Jenkins, J.P., Braswell, B.H., Hollinger, D.Y., Ollinger, S.V., Smith, M.L. 2007. Use of digital webcam images to track spring green-up in a deciduous broadleaf forest. Oecologia 152, 323–334.
- Roth-Nebelisck A., Uhl D, Mosbrugger V., Kerp H. 2001. Evolution and function of leaf venation architecture: a review. Annals of Botany, 87: 553-566.
- Sack L., Scoffoni C., McKown A.D., Frole K., Rawls M., Havran J.C., Tran H., Tran T. 2012. Developmentally based scaling of leaf venation architecture explains global ecological patterns. Nature communications. DOI: 10.1038/ncomms1835.
- Sack L., Scoffoni C. 2013. Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 198: 983-1000.
- Sonnentag O., Hufkens K., Teshera-Sterne C., Young A. M., Friedl m., Braswell B. H., Milliman T., O’Keefe J., Richardson A. D. 2012. Digital repeat photography for phenological research in forest ecosystems. Agricultural and Forest Meteorology. Vol. 152, pp. 159-177.
- Woebbecke D.M., Meyer G.E., Vonbargen K., Mortensen D.A. 1995. Color indexes for weed identification under various soil, residue, and lighting conditions. Transactions of the ASAE, 38: 259–269.
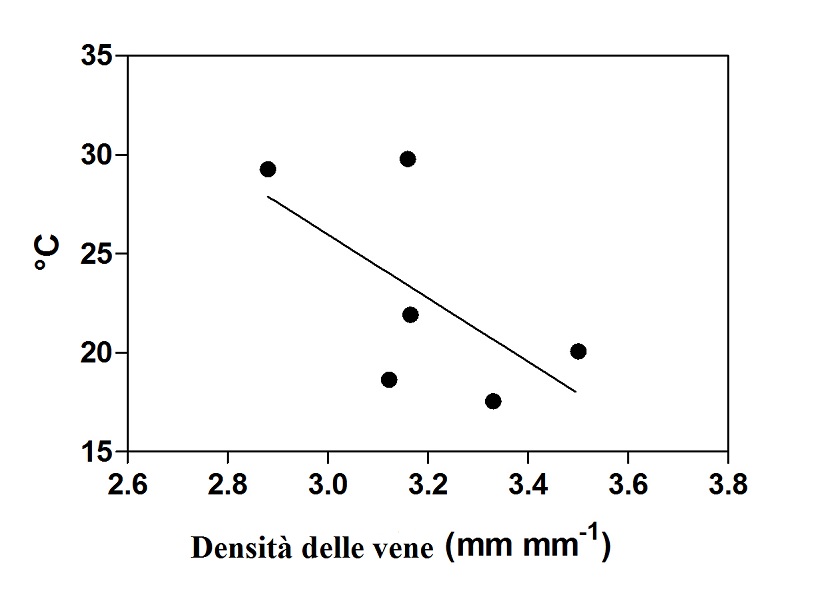
Fig. 1 – Correlazione significativa tra la densità delle vene e la temperatura della foglia (P< 0.01, r² = 0.39, no.= 6)(Pagano et al., 2015).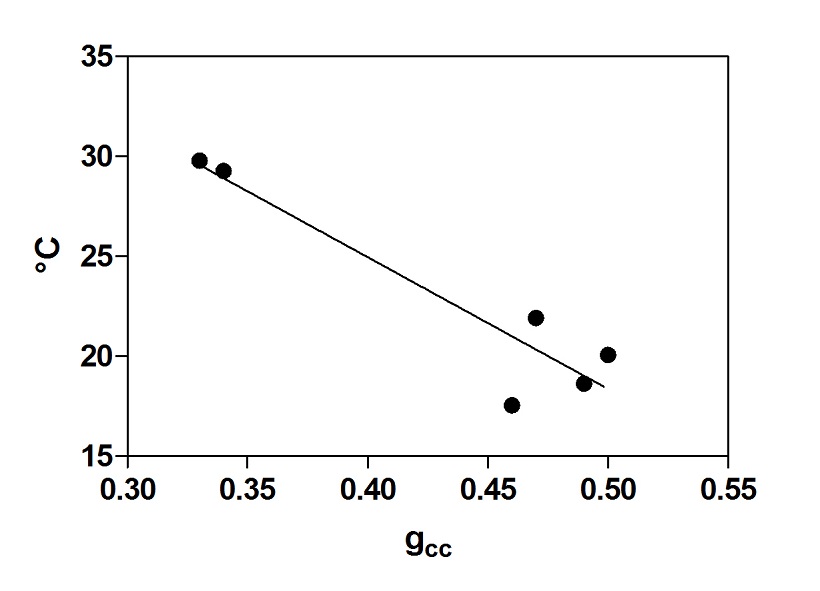
Fig. 2 – Correlazione significativa tra l’indice gcc e la temperatura della foglia (P< 0.01, r² = 0.86, no.= 6) (Pagano et al., 2015).
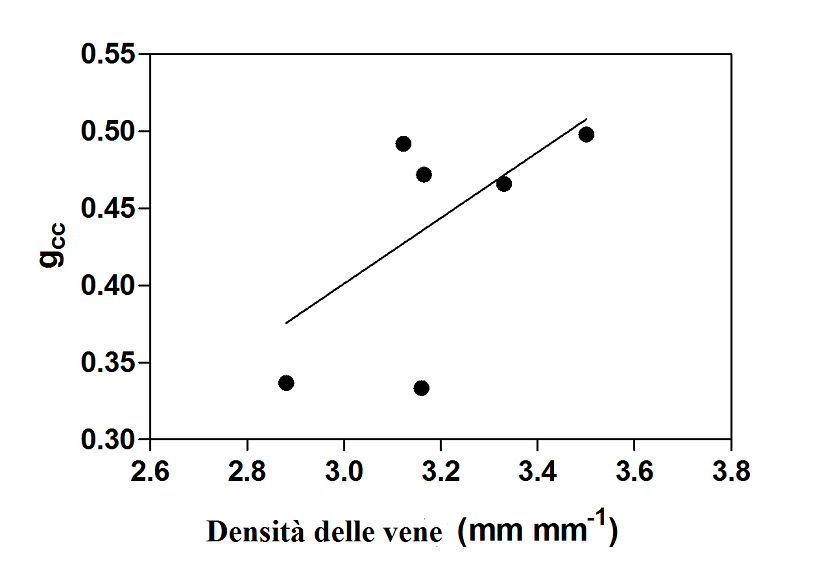
Fig. 3 – Correlazione significativa tra l’indice gcc e la densità delle vene (P< 0.01, r² = 0.36, no.= 6) (Pagano et al., 2015).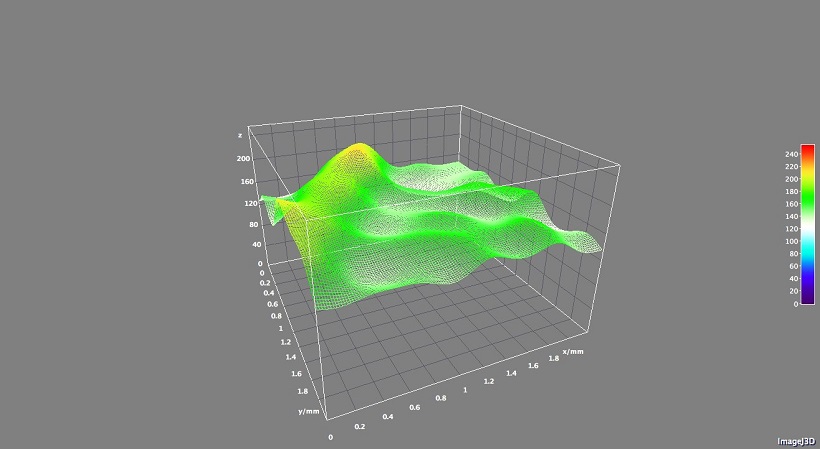 Fig. 4 – Esempio di spettro termico virtuale (LUT) di una porzione di lamina della foglia (Pagano et al., 2015).
Fig. 4 – Esempio di spettro termico virtuale (LUT) di una porzione di lamina della foglia (Pagano et al., 2015).
Mario Pagano, laureato in Scienze e Tecnologie Agrarie (vecchio ordinamento) presso l’Università degli Studi di Firenze, ha conseguito il Dottorato di ricerca in Agrobiotecnologie per le Produzioni Tropicali. Curriculum vitae >>>
|






