
di Pierluigi Simonini
Premessa
Questo articolo rappresenta una sintesi di quanto ho letto sulla genetica delle cavie nel corso degli ultimi due anni.
La bibliografia di riferimento non è molto ampia, ma è abbastanza completa di contenuti da consentirmi di muovere i primi passi nei principi base che regolano la riproduzione delle diverse razze e colori delle cavie di selezione.
I concetti espressi negli articoli a cui ho fatto riferimento, non erano sempre coincidenti, e inevitabilmente sono stato influenzato più da alcuni che da altri, in questi casi, per quanto nelle mie possibilità, ho sempre cercato di orientarmi tenendo conto o di esperienze personali o di quelle riferitemi da altri allevatori.
Chi ha scritto questo articolo non è né un medico, né tanto meno un veterinario o un esperto di genetica, ma solo un appassionato, incuriosito da tutti quegli aspetti estremamente importanti della genetica di base delle cavie, la cui conoscenza e comprensione è indispensabile nella selezione di questi piccoli animali.
Leggendo quanto scritto nell’articolo, talvolta potrà sembrare che vi sia la presunzione di poter racchiudere in poche tabelle e pochi sintetici concetti delle verità assolute, ma vi posso assicurare che non è così.
Dobbiamo tenere sempre in considerazione, che a quanto pare, vi sono moltissimi locus genici riferiti alle cavie, e che noi ne abbiamo illustrati e presi in considerazione poco più di una decina.
Ci rendiamo immediatamente conto, che molte cose o ci sfuggono o le possiamo solo immaginare e teorizzare, e che in presenza di una tale percentuale di incognite, che oltretutto possono interagire fra loro, parlare di verità e sfoderare concetti sicuri è quanto di più lontano dalla realtà ci possa essere.
I locus genici che siamo riusciti a prendere in considerazione, anche se sono solo poco più di una decina, in ordine d’importanza sono sicuramente i più rappresentativi, ma non perché lo pensiamo noi arbitrariamente, piuttosto perché sono frutto di riscontri oggettivi della realtà delle cose e rappresentano al momento le risposte più corrette che esperti di genetica e ricercatori hanno dato a molte delle nostre domande.
Seppur in un quadro d’incertezza complessiva alimentato continuamente da dubbi e domande, quanto illustrato di seguito, rappresenta una solida ossatura attorno alla quale possono trovare una corretta collocazione tutti i concetti e le conoscenze che ancora ci sfuggono e non conosciamo.
Nella parte iniziale di questo articolo, si è resa necessaria una brevissima introduzione ai concetti e alle terminologie, che si sarebbero trovati di lì a poco, nell’illustrazione dei concetti base della genetica dei colori.
Nella speranza di non avere commesso troppi errori, buona lettura.
Breve introduzione alla genetica
Geni&Alleli
Ogni carattere ereditario si trova sotto il controllo di una coppia di fattori detti Geni.
Le differenti caratteristiche che può assumere lo stesso gene si chiamano Alleli (X;x).
Tutti gli individui possiedono una singola coppia di alleli per ogni Carattere ereditario (ad es.B;b).
Quando la coppia responsabile di un carattere è formata da alleli identici (XX; xx) l’individuo è detto “omozigote” per quel determinato carattere.
Ogni singolo sito che ospita la coppia di alleli è detto locus genico.
Quando la coppia è formata da alleli diversi l’individuo è detto “eterozigote” (Xx) per quel determinato carattere.
Ogni coppia di alleli è responsabile ciascuna di un determinato carattere ereditario: colore degli occhi, del mantello (nel particolare caso delle cavie, ecc.).
In una singola coppia dove gli alleli sono diversi, l’azione di ciascun allele può essere dominante (X-) o recessiva (xx).
In queste coppie con alleli diversi, quella più “forte” che si manifesta nell’individuo viene detta dominante, mentre quella che non si manifesta è detta recessiva.
Quindi nei cromosomi si trovano tutte le informazioni ereditarie che permettono la “costruzione” dell’individuo.
Il gene è dunque una parte di cromosoma che descrive una certa caratteristica di quell’individuo.
Una o più singole coppie di alleli possono influenzare, modificare l’effetto di una o altre coppie o rendere non espresse nel fenotipo la loro azione, queste coppie rimangono latenti ed impossibilitate ad esprimersi sino a quando non vi sono nella prole condizioni mutate del genotipo che consentono la loro espressione.
I caratteri di un individuo sono descritti da molti geni, e molte volte un carattere dipende da più di un gene, l’insieme dei geni di un individuo è detto Genotipo.
Quindi il genotipo è tutto quello che si trova nei cromosomi.
L’insieme dei caratteri di un individuo è detto fenotipo, che rappresenta tutto ciò che possiamo osservare di un individuo e che non sempre coincide col genotipo.
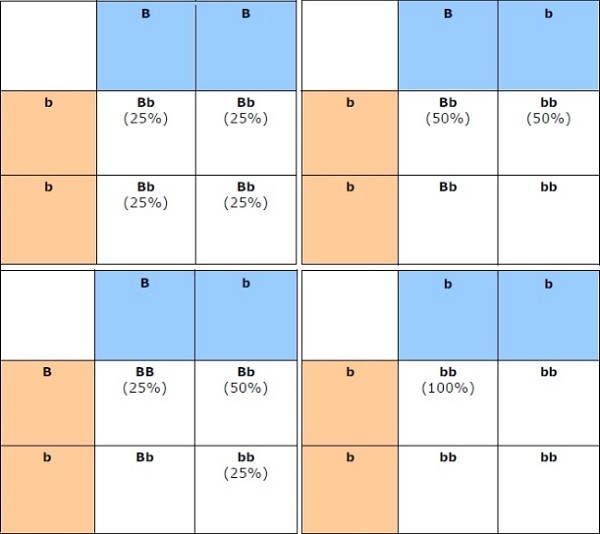
Per spiegare meglio quanto e cosa può essere trasmesso alla prole e le percentuali teoriche che ciascuno appartenete a questa prole abbia ricevuto determinate caratteristiche ci viene in aiuto il Quadro di Punnet.
Noi dobbiamo immaginare di ripetere quanto riportato come esempio nei differenti quadri di Punnet indicati qui di seguito, come rappresentazione delle varie possibilità di trasmissione dei caratteri alla prole di prima generazione (F1), per ogni singola coppia di alleli, di ogni singolo Locus Genico della sequenza del genotipo delle due cavie “padre” e “madre”, è ovvio che più coppie di alleli conosciamo e prendiamo in considerazione dal genotipo dei genitori, più le variabili e le possibili combinazioni viste nel loro complesso si moltiplicano.
Nei seguenti quadri di Punnet, prendiamo in considerazione ad esempio il solo locus genico degli alleli del gene del colore Nero (B-) e quello del suo subordinato e recessivo gene del colore Cioko (bb).
In azzurro abbiamo il locus genico del padre (parte orizzontale della tabella), e in rosa quello della madre
(parte verticale laterale sx), incrociando i singoli alleli di entrambi nella tabella otteniamo le possibili caratteristiche ricevute nella loro prole, dove “teoricamente” ogni riquadro rappresenta il 25% delle possibilità che ciò accada .
Gli allevatori sanno bene che queste percentuali sono solo “teoriche”, o meglio: sono veritiere se riferite ai “grandi numeri”, ad esempio se lancio una moneta 5000 volte molto probabilmente alla fine avrò ottenuto più o meno per metà delle volte testa e per metà delle volte croce, ma se la lancio solo una decina di volte sarà tutto più casuale, posso anche ottenere quasi sempre testa e quasi mai croce.
In quest’ottica dico che se faccio una sola o poche volte l’accoppiamento del nostro terzo caso, le possibilità possono rispecchiare le probabilità di ciascun colore nella prole, ma potrei ottenere anche cucciolate in cui quello che doveva risultare solo per il 25% delle possibilità di fatto potrebbe essere anche l’80/100%, quindi se ci riferiamo ai “piccoli numeri” diciamo che è più veritiero considerare quando risulta dal quadro di Punnet solo per i colori possibili nella prole e non tanto per le loro percentuali.
· • Primo caso : Padre Nero omozigote (BB), Madre Cioko (bb)
· • Secondo caso : Padre Nero eterozigote (Bb, carrier cioko), Madre Cioko (bb)
· • Terzo caso : Padre e Madre Nero eterozigote (Bb)
· • Quarto caso : Padre e madre Cioko (bb, essendo recessivo per esprimersi deve essere bb)
Quadro di Punnet
La Genetica dei colori nelle Cavie
Il Nero&il Rosso
Generalizzando un po’ possiamo dire che per quel che riguarda i colori del mantello delle cavie, ci troviamo di fronte a due soli colori:
1. Nero B
2. Rosso ee
questa affermazione non è del tutto corretta, o meglio: ci troviamo in presenza di un solo colore che potremmo definire principale o “dominante” il Nero con i suoi alleli, e un serie un gruppo di alleli appartenenti ad una medesima “famiglia” di geni, che in modo progressivo consentono l’espressione ed il manifestarsi del colore Rosso.
Più precisamente, consentono al colore Nero di estendersi e “coprire” quello Rosso fino a farlo scomparire.
Questi alleli detti di “estensione”, appartengono alla “famiglia” del gene E , vedremo il loro modo di comportarsi qui di seguito.
Prima di fare ciò, è necessario integrare il discorso con una precisazione: il colore Nero (B-) può essere modificato dal gene b, responsabile di una modificazione del pigmento nero del mantello, quando b può esprimersi in omozigosi (bb) il colore Nero viene ad assumere una colorazione color cioccolato, o più precisamente, il color Cioko.
L’allele b però è più “debole” del B suo dominante, infatti, mentre B si presenta come “dominante” e come tale si comporta (perché sia manifesto è sufficiente che sia presente anche solo una volta in eterozigosi), il suo subalterno b ha caratteristiche più “deboli “ di tipo “recessivo”,e per prevalere su B e rendere manifesti i suoi effetti, deve scalzarlo del tutto ed occupare entrambi i posti disponibili, presentandosi congiuntamente a se stesso come (bb), per così dire in omozigosi.
La gamma di possibilità potrà essere la seguente:
1. BB Colore Nero, B in omozigosi, tutta la prole Nera
2. Bb Colore Nero, B in eterozigosi, Nero portatore di Cioko, prole o Nera o Cioko a seconda delle caratteristiche del compagno
3. bb Colore Cioko, b in omozigosi, prole Nera o Cioko a seconda del compagno, ma sicuramente tutta portatrice di Cioko
Ora torniamo ai geni della “famiglia E detti di “estensione”, questi geni che impropriamente possiamo chiamare del colore Rosso sono i seguenti:
1. E Totale estensione della componente Nera o Cioko
2. ep Totale estensione della componente Tortie o Brindle
3. e Totale estensione della componente Rossa
In questo medesimo ordine hanno, a scalare, caratteristiche di dominanza sul subalterno sottostante, o meglio: E sarà dominante su ep e su e, mentre ep sarà dominante solo su e e subalterno a E, mentre e non è dominante su nessuno degli altri due e sarà il più recessivo di tutti e come tale per esprimersi
dovrà, per forza di cose, presentarsi abbinato al suo omologo, ovvero secondo la sequenza: ee che così com’è, rappresenta il colore Rosso.
Quindi, secondo quanto detto, possiamo avere i seguenti abbinamenti nel locus di E:
1. EE Espressione totale della componente Nera o Cioko del mantello
2. Eep Espressione totale della componente Nera o Cioko, ma come portatore di tortie
3. Ee Espressione totale della componente Nera o Cioko, ma come portatore di rosso
4. epep Espressione totale della componente Tortie o Brindle
5. epe Espressione totale della componente Tortie o Brindle, ma come portatore di rosso
6. ee Espressione totale della componente Rossa
Se noi consideriamo il colore Nero ed il colore Rosso come i principali colori che originano tutta un’infinità di altri colori, ci dobbiamo anche occupare di come, ed in che modo, tutto questo possa avvenire.
Cercherò di spiegare per quanto mi è possibile in modo chiaro e sintetico, quali sono i meccanismi che consentono che ciò avvenga.
Generalizzando un po’ possiamo dire che abbiamo due meccanismi di modificazione o diluizione del colore Nero e del colore Rosso del mantello delle cavie.
Diluizioni e modificazioni del Nero e del Rosso
1. I geni della “famiglia” P che originano le varietà dei colori negli occhi
2. I geni della “famiglia” C che modificano il colore Rosso
I geni della famiglia P : quali il P o il pg, oppure il p, determinano rispettivamente un occhio Nero, un occhio Rubino, o un occhio Rosa, questo gruppo di alleli hanno una marcata azione nella scoloritura del colore Nero del mantello e agiscono rispettivamente così: il Nero rimane inalterato quando siamo in presenza di P (occhio nero), il Nero diventa Slate quando abbiamo in omozigosi l’allele pg (pgpg, occhio rubino), quindi possiamo dire che il colore Slate è uguale ad un Nero con occhi rubino (abbiamo uno scolorimento del pigmento che può arrivare anche fino al 50% ), il Nero si trasforma in lilla quando al colore Nero si unisce in omozigosi l’allele p (pp, occhio pink), quindi possiamo dire che il colore Lilla è uguale ad un Nero con occhi pink (scolorimento del mantello fino l’80% del suo colore).
Tutti gli alleli di questa “famiglia”, che, come abbiamo visto, esplicano un’azione molto incisiva di scoloritura del colore Nero, hanno un effetto però solo limitato e marginale sulla componente Rossa. Però diventano una concausa di quelle sfumature più o meno percepibili dell’intensità del colore, che possono fare la differenza fra un soggetto che noi ad esempio reputiamo colorato in modo più corretto piuttosto che un altro.
Diversamente dall’azione dei geni P, tutti i geni della “famiglia” C hanno un’azione solo marginale di scoloritura del colore Nero, ma progressivamente anche molto intensa del colore Rosso.
Appartengono a questa “famiglia”, il C che genera il Rosso nella sua essenza più scura, il cd[*] che rappresenta un primo modesto grado di scoloritura e in omozigosi (cd cd) origina il colore Buff, e altri
due, quali il cr ed il ch, che congiunti al cd intensificano l’opera di scoloritura del colore rosso a Crema.
[*] (cd): ha due effetti collaterali alla sua azione di diluizione del Rosso: provoca una lievissima scoloritura del pigmento nero del pelo e una modesta scoloritura dell’occhio nero (l’occhio rimane comunque nero nel suo genotipo).
(ck): è stato menzionato un altro allele ck di scoloritura del rosso capace di originare un colore Buff scuro, ma ad oggi la sua presenza e il suo meccanismo d’azione e oggetto di osservazioni e discussioni, sicuramente l’azione dell’allele cd e l’interazione di altri fattori ad influenza minima è più che sufficiente a spiegare le possibili colorazioni e sfumature in particolare del Buff.
(cr): ha due effetti collaterali come cd: provoca una leggera scoloritura del pigmento nero del pelo e porta l’occhio nero ad una tonalità rubino scuro (l’occhio rimane comunque nero nel suo genotipo) (c) esiste anche un allele c ma è molto raro, per questo motivo per ora non lo prendiamo in considerazione, è di tipo recessivo, è il gene dell’albinismo, causa una depigmentazione totale e particolari problemi alla vista.
Il cr può essere definito un allele di intensa scoloritura del rosso, così come il ch, che è più propriamente l’allele del colore himalayan , entrambi hanno la capacità di portare il colore Rosso a Bianco.
L’allele ch in omozigosi (chch) è responsabile proprio del colore Himalayan.
Gli alleli cr e ch, si possono presentare anche in modo disgiunto rispetto a cd, possiamo avere la sequenza (cr cr) che genera il mantello completamente bianco, ma solo quando la cavia è “genotipicamente Rossa”(ee), oppure (cr ch), sempre col medesimo risultato se a parità di condizioni (ee), ma diversamente dalla precedente, questa cavia bianca sarà portatrice del colore himalayan proprio perché ottenuta con l’aiuto del gene ch.
La sequenza (ch ch), sempre a parità di condizioni (ee), genera il medesimo risultato, abbiamo sempre un mantello totalmente bianco, ma questa volta con l’occhio di colore rosa, quale caratteristica a corredo proprio della colorazione himalayan.
In questo caso possiamo dire che la cavia sarà un himalayan di “base” rossa, quindi tutta bianca, anche alle estremità, sempre come espressione del fatto che il gene himalayan ha un’azione di scoloritura totale sul colore rosso.
In quest’ultimo caso, se avessimo voluto ottenere una cavia himalayan dalla colorazione corretta e le estremità nere o al più cioko, alla sequenza (ch ch) dovevamo unire l’azione esercitata dalla sequenza (E-) che consente solo l’espressione del colore Nero o Cioko e “sopprime” il colore Rosso.
Tutti i geni della “famiglia” C che come abbiamo visto, possono esercitare una forte azione di scoloritura del Rosso, ed essere quasi ininfluenti sul Nero, influenzano leggermente anche la pigmentazione dell’occhio, contribuendo alle varie sfumature che un occhio di un determinato colore può avere, come avviene ad esempio per l’occhio “fuoco”.
L’occhio fuoco infatti non viene generato da un suo specifico singolo gene dell’occhio fuoco, ma dagli “effetti collaterali” esercitati dai geni della famiglia C, ora su un occhio rubino, ora su un occhi nero, questi effetti possono rendersi anche manifesti in una colorazione non proprio uniforme delle setole del mantello dei Neri o dei Cioko.
Se a questi effetti “collaterali”, anche se modesti, uniamo altrettanti effetti modesti, che gli alleli della “famiglia” P esercitano sul colore Rosso, e a questo aggiungiamo le azioni più marcate che sia P che C hanno nelle loro rispettive aree di competenza (C per il Rosso e P per il Nero), ci rendiamo immediatamente conto di quante possibili sfumature e variazioni di colore del mantello possiamo ottenere, anche se in presenza di due sole “variabili base”: il Nero ed il Rosso.
Congiuntamente a questo immaginiamo quante ulteriori casistiche possiamo avere se a questi effetti uniamo l’azione di altre importanti famiglie di geni, quali ad esempio: gli Agouti, i Pezzati di bianco aprendo ai tricolori, Dalmata e Roani e mix vari fra tutti questi geni ecc.
E pensare che stiamo parlando solo di colorazioni del mantello, ora proviamo ad immaginare di unire a questa moltitudine di combinazioni e possibili colori, tutta una serie di geni collegati alla lunghezza del mantello del pelo (corto, semi-lungo, lungo), alla sua consistenza per durezza ruvidità ecc. famiglie dei geni Rexoid o Us teddy o Ch teddy ecc.), al modo di presentarsi e piegarsi (famiglie dei geni delle rosette e tutti i loro soppressori e regolatori anche parziali).
Questa moltitudine di geni, anche solo teoricamente, potrebbero essere mescolati in una serie di combinazioni che come potrete immaginare originerebbe una miriade di morfologie, colori o razze.
E’ stato calcolato che sarebbe possibile ottenere oltre 280 “razze” di cavie mescolando tutte queste variabili, magari alcune esteriormente molto simili, ma ottenute da combinazioni di geni molto dissimili fra loro, in quest’ottica ci si rende immediatamente conto della necessità di inquadrare più rigidamente un numero contenuto di “razze”.
Questo è stato il percorso perseguito dalle varie associazioni e dai tanti appassionati allevatori, che hanno contribuito ad inquadrare in anni di selezione molte delle razze oggi conosciute ed apprezzate per la loro qualità.
Noi per ora, con questo articolo rimaniamo nell’ambito delle sole colorazioni possibili, ed impariamo a conoscere e a prendere dimestichezza con i geni che le regolano, presto ci renderemo conto
dell’importanza di sapere scrivere le sequenze dei genotipi delle nostre cavie, magari cercando d’integrare i dati a noi mancanti con quelli ottenuti anche solo da semplice ragionamento osservando le colorazioni dei loro figli.
E’ importante avere sempre sott’occhio e leggere le tabelle delle sequenze dei colori (genotipi dei colori o “riconosciuti” o “possibili”), queste tabelle rappresentano un aiuto alla sintesi ed alla comprensione dei colori stessi e del come ottenerli dai vari accoppiamenti, ma saranno utili soprattutto per comprendere il “perché” di un colore più che il suo inquadramento in uno standard espositivo.
Tabella dei genotipi delle principali colorazioni self non agouti
Con quanto detto sino ad ora, siamo in grado di capire almeno le sequenze base dei genotipi delle principali colorazioni Self, prive delle “contaminazioni” di tutta quella serie di geni che vedremo in seguito e che moltiplicheranno inevitabilmente la gamma dei possibili colori del mantello.

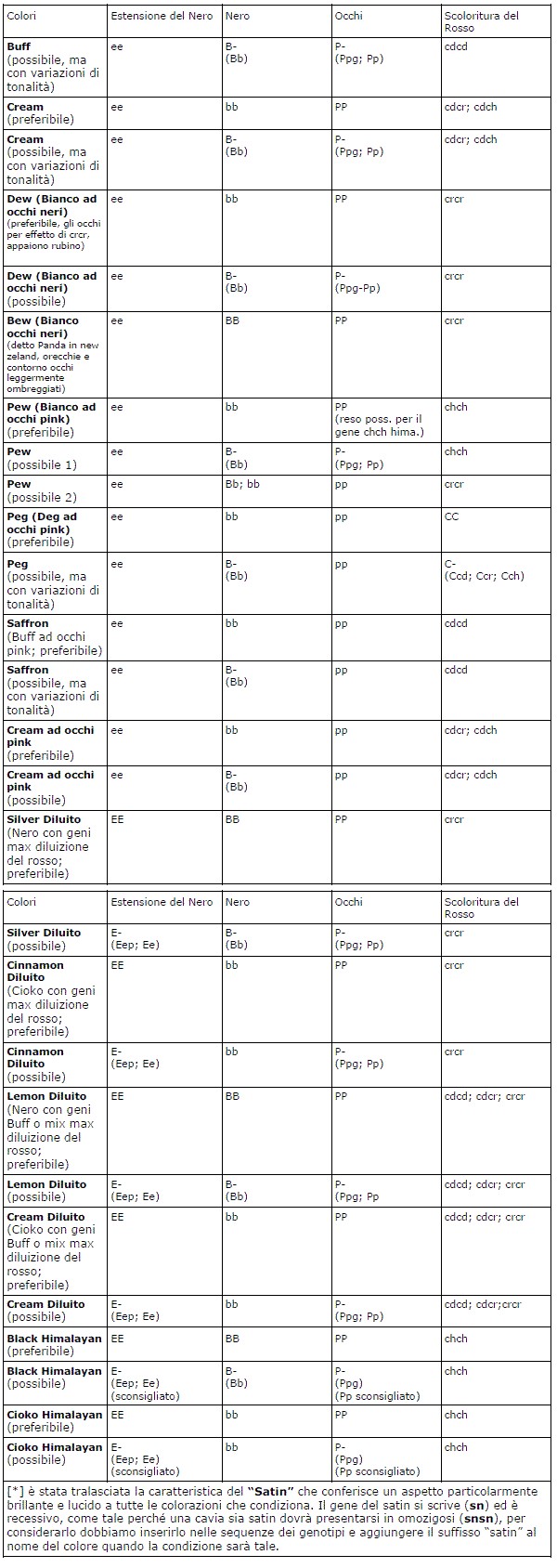
In questa tabella, che riassume le sequenze possibili dei principali colori self (non pezzati di bianco e nonagouti), ho inserito in voci distinte il concetto di “preferibile” e quello di “possibile”.
Per “preferibile” si intendono le condizioni ideali di “purezza”della sequenza del genotipo che consentono, in quanto tali, di manifestare ed esprimere al meglio le caratteristiche di ogni singolo colore.
Avrete notato che ciò avviene quanto più accade che la “voce” di ogni singolo gene si esprime in
omozigosi, anche quando non sarebbe stato necessario per manifestarne gli effetti, come ad esempio per i geni dominanti.
Tutto questo è giustificato dal fatto che la condizione di omozigosi di un allele anche se dominante, di fatto, non consente ad un omologo recessivo di occupare l’altro posto eventualmente disponibile nel particolare locus genico.
Questi alleli recessivi, pur non esprimendosi perché sotto l’effetto del loro dominante, talvolta riescono ad avere ugualmente una minima influenza sulla colorazione del mantello o degli occhi, magari quasi impercettibile, ma che se abbinata ad altrettante azioni quasi impercettibili di altri alleli recessivi, contribuiscono complessivamente ad incrementare o a sbiadire una tonalità e rendere meno uniforme una colorazione.
Per questo motivo, è sempre “preferibile” avere quanti più alleli dominanti in condizione di omozigosi nei rispettivi locus delle sequenze dei genotipi dei vari colori.
Spesso allevatori impegnati nella selezione di determinati colori, “lavorano” esclusivamente su uno solo, o max. due colori, abbinando sempre colori simili e per quanto possibile vicini allo standard, inserendo nel programma di allevamento preferibilmente soggetti anch’essi frutto di un identico criterio di selezione.
Questo tipo di programma, rappresenta quasi una necessità per chi intende avere il minor numero di soggetti con colorazioni non ottimali e meno “sorprese” in ambito di selezione.
Ovviamente soggetti con un genotipo di questo tipo, pur rappresentando la condizione ottimale, spesso fatichiamo a reperirli o ad avere la certezza che il loro genotipo abbia veramente queste determinate caratteristiche.
E’ in quest’ottica, che la conoscenza genetica del perché di un colore e del come un medesimo colore può essere frutto di genotipi diversi, ma esprimere fenotipi quasi indistinguibili, ci viene in aiuto, e diventa una conoscenza indispensabile per fare convergere il nostro lavoro di selezione verso un soggetto con caratteristiche ottimali.
La maggior parte dei soggetti che abbiamo ricadono nella condizione di colore “possibile” o meglio di una colorazione anche perfetta, ma non proprio al 100% delle proprie possibilità, da inserire in ambito riproduttivo con cognizione di causa, cercando di evitare accoppiamenti con soggetti che presentano delle eterozigosi possibili causa di sorprese non del tutto gradite nella prole.
Una volta compreso questo meccanismo, nelle prossime tabelle sarò più sintetico, indicando con più frequenza esclusivamente genotipi di tipo ottimale o il solo riferimento all’allele dominante (X-) , anche perché come avrete capito, l’elenco delle possibili variabili renderebbe troppo “gonfie” le tabelle che al contrario dovrebbero essere espressione di sintesi.
Indicherò sempre nei genotipi o l’allele dominante in omozigosi (XX), o la sua possibile “apertura” ad un generico suo omologo recessivo (X-), lasciando sottinteso che lo stesso colore si può ottenere in minime variazioni di tonalità con la presenza dei rispettivi alleli recessivi.
Rimane il fatto che, non avendo io una conoscenza approfondita dei colori riconosciuti dalle varie associazioni, parlando in termini espositivi, e in considerazione del fatto che un medesimo colore viene chiamato ed indicato a volte con nomi diversi, riporterò i nomi dei colori utilizzando le diciture che ho trovato all’interno della mia specifica bibliografia di riferimento a questo articolo.
Sono sicuro, che ove si presentasse questo inconveniente, sarà comunque di facile comprensione capire lo specifico colore a cui mi riferisco.
Pierluigi Simonini – Vice presidente ANAC
![]()
L’ANAC promuove la diffusione delle cavie domestiche (Cavia porcellus), incoraggia l’allevamento, lo studio e il miglioramento delle diverse razze, ai fini di incrementarne la conoscenza e la valorizzazione come animali sociali e d’affezione.
http://www.agraria.org/cavie/anac.htm
 |
Il Coniglio nano – Agraria.org Cristiano Papeschi – Il Sextante In modo chiaro e simpatico vengono trattati tutti gli aspetti legati alla sua presenza in casa, dal ricovero all’alimentazione, dalla convivenza con gli altri animali alle cure quotidiane, dalla riproduzione alla sua salute. Per ricevere una copia scrivere a: info@agraria.org |



")

