
di Umberto Mazzucchi
Le tossinfezioni alimentari possono essere causate da infezioni di microrganismi patogeni associati alle mucose intestinali oppure alla ingestione di alimenti da loro contaminati o da tossine microbiche anche in assenza di microrganismi produttori. Sono note circa 250 forme di tossinfezioni alimentari e almeno 10 specie batteriche agenti potenziali, tra cui Salmonella enterica e forme tossigeniche di Escherichia coli hanno maggiore notorietà. Nel 2010, nella Unione Europea, le diagnosi di 27 casi su 70 di zoonosi causate da consumo di frutta e verdura hanno identificato batteri come agenti causali (1); infezioni di Salmonella sono state accertate in 8,6% dei casi. Nell’ aggiornamento epidemiologico 2010 delle malattie trasmesse da alimenti, la Regione Emilia Romagna ha segnalato che 36 casi su 89 sono stati attribuiti a ceppi di S.enterica, nessuno a forme tossigeniche di E. coli. Com’è noto, hanno suscitato allarme nella opinione pubblica e interventi delle autorità sanitarie le epidemie scoppiate nel 2011 in Germania e in altri paesi europei, attribuite dapprima al consumo di cetrioli, poi a quello dei germogli di leguminose (2). Esperti prevedono che in futuro ci possa essere incremento di tossinfezioni da consumo fresco di vegetali.
In passato si credeva che l’ associazione di batteri zoopatogeni a prodotti vegetali fosse conseguenza di mera contaminazione occasionale di superficie in pre- o – in post –raccolta o al momento del consumo, causata da scarsa igiene dell’ambiente e degli operatori. Nell’ultimo decennio microbiologi hanno messo in luce che sia S.enterica che le forme tossigeniche di E.coli non solo riescono a penetrare nei tessuti vegetali, ma possono moltiplicarsi al loro interno (stato endofita) e colonizzare la pianta anche a distanza dai punti di penetrazione. In piante di pomodoro, ad esempio, S.enterica inoculata nei fusti può raggiungere i tessuti interni dei frutti, sia pure con bassa frequenza (3). Lo stato endofita accertato per i due batteri e in futuro verosimilmente anche per altri batteri zoopatogeni è inevitabile conseguenza sia della penetrazione passiva dei batteri negli organi vegetali, sia del tipo di risposta del sistema immunitario delle piante ai procarioti pervenuti in contatto con i tessuti interni.
Per quanto riguarda la penetrazione, secondo la teoria della congestione idrica, nota in patologia vegetale dalla metà del secolo scorso, qualsiasi batterio sospeso nel liquido di bagnatura di certe sedi può penetrare passivamente all’interno di organi vegetali in condizioni di eccesso di acqua nella pianta. Sono sedi di penetrazione le aperture naturali (es. stomi, pori degli idatodi e dei nettari, spazi intercellulari degli stigmi) e le micro- e macro-ferite (es. rottura di tricomi alla base, screpolature nel tessuto di riempimento delle lenticelle, erosioni o punture di fitofagi, ferite da grandine, etc.). Creatasi una continuità temporanea (bastano pochi minuti) tra i liquidi interni e il liquido di bagnatura (rugiada, pioggia, irrigazione), le cellule batteriche contaminanti il bordo della sede sono aspirate verso l’interno della colonna liquida che arretra per effetto della evaporazione. Più spesso la penetrazione ha luogo in campo durante la stagione vegetativa alla levata del sole in presenza della bagnatura di rugiada quando si riaprono gli stomi ed è finita la reidratazione della notte. Essendo la penetrazione passiva, qualsiasi batterio può pertanto essere introdotto all’interno di tessuti vegetali, incluse specie zoopatogene, se per caso vengono a trovarsi nei liquidi di bagnatura di foglie o frutti. Batteri mobili per mezzo di flagelli, tuttavia, possono attivamente accelerare l’ingresso nella pianta. Non sempre i batteri penetrati riescono a colonizzare i tessuti in profondità. In certi casi si ammette che le loro popolazioni vengano a trovarsi in microscopici rifugi immediatamente sotto la superficie (nicchie semi-interne) come, ad esempio, spazi intercellulari ampi sotto le aperture degli stomi (camere sottostomatiche) o degli idatodi o entro le fessure microscopiche associate a micro-ferite di rottura di tricomi nelle foglie, negli spazi tra le cellule del tessuto di riempimento delle lenticelle nei frutti o nei tuberi. Queste nicchie interne o semi-interne offrono sempre ai batteri un certo grado di protezione all’azione degli interventi igienici, siano lavaggi o applicazioni di mezzi fisici o chimici.
Per quanto riguarda la risposta del sistema immunitario della pianta, si sa che la cellula vegetale venuta in contatto di parete con un batterio penetrato dà luogo entro 3-4 ore a una risposta localizzata espressa sotto forma di barriere fisico-chimiche antibatteriche o meglio antimicrobiche; contemporaneamente anche il batterio esprime risposte per cercare di adattarsi alla nuova nicchia. Sono ore cruciali per il destino del batterio nel tessuto. In realtà questa risposta tempestiva ha luogo tutte le volte che una cellula vegetale viene in contatto con una entità che riconosce estranea a sé stessa (riconoscimento del non-self). Studi sulle interazioni piante-batteri hanno mostrato che nei confronti di una entità vivente non-self presente nell’apoplasto le cellule vegetali attuano un riconoscimento in negativo: tollerano l’estraneo, probabilmente quasi per saggiare la possibilità di integrazione, fino a quando l’estraneo non trasmette un segnale (-i) di incompatibilità È una strategia vincente che nel corso dell’evoluzione dà alla pianta maggiore probabilità di creare simbiosi mutualistiche. Questo comportamento spiega molto bene la molto frequente presenza di microrganismi all’interno di organi vegetali, più spesso asintomatici; in particolare di procarioti, adattatisi alla vita endofita tanto da permanere tali anche per tutta la vita di piante poli-annuali. La numerosità delle loro popolazioni viventi endofite rimane controllata permanentemente da strumenti del sistema immunitario della pianta. Più lunga e ripetuta è la permanenza endofita, tanto maggiori sono tempo di selezione e probabilità di adattamento. I batteri endofiti patogeni virulenti per una pianta esprimono invece fattori che trasferiti con sistemi di secrezione nelle cellule vegetali interferiscono con la risposta immunitaria e la menomano in modo da potersi moltiplicare agevolmente e colonizzare progressivamente nuovo tessuto durante il così detto periodo di incubazione. In accordo alla teoria della risposta quantitativa (4), anche per i patogeni virulenti, tuttavia, si giunge a un punto della interazione (es. maculature) dove la intensità della risposta delle cellule vegetali alla attività del microrganismo endofita diviene tale da non essere più compatibile con la omeostasi dei suoi processi metabolici e si avvia un processo degenerativo espresso dalla comparsa dei sintomi tipici di malattia.
Tenuto conto di questi tipi di rapporti intercorrenti tra batteri e piante, è facile comprendere come anche batteri zoopatogeni possano penetrare all’interno di organi vegetali e svolgere vita endofita, moltiplicandosi, soprattutto se introdotti sperimentalmente ad alte dosi nell’apoplasto. La loro presenza associata a tossinfezioni dell’uomo conseguenti al consumo di vegetali freschi è stata interpretata attribuendo alle specie vegetali coinvolte il ruolo di piante ospiti alternative (5). Secondo questa interpretazione, ospite principale sarebbe l’uomo soggetto alle tossinfezioni, ospiti secondari animali domestici o selvatici (es. bovini, polli, maiali…cervi, passeri…) e piante (es. specie vegetali da consumo fresco), mentre sorgenti di inoculo sarebbero residui di escrementi animali (es. letame), terreno, acque di irrigazione e vegetali consumati freschi. Questa interpretazione deve essere accettata con cautela. Per attribuire ad una specie vegetale il ruolo di pianta ospite per un batterio, i protocolli della batteriologia fitopatologica raccomandano di usare basse dosi di inoculo, vie di penetrazione naturali e di applicarle su piante aventi stadi fenologici assimilabili a quelli di campo, in condizioni termo-igrometriche appropriate. Gli studi sulla endofitia dei batteri zoopatogeni non sempre sono state conformi ai canoni della fitobatteriologia (6). D’altra parte solo recentemente è stato messo in luce che i ceppi trovati nelle piante possono essere differenti da quelli causanti le infezioni nell’uomo, pur essendo filogeneticamente affini (7). La diversificazione è verosimilmente conseguenza del loro adattamento da un lato all’ambiente endofita, dall’altro lato agli stress ambientali connessi alla loro inevitabile, temporanea permanenza in acqua, nel terreno o nella fillosfera prima di penetrare nella pianta. Se la pressione selettiva multipla cui sono stati sottoposti si è tradotta in modificazioni genomiche, certi caratteri possono essere stati fissati nelle popolazioni e trasmessi verticalmente. Così, ad esempio, una alta frequenza di ceppi di E. coli isolati da vegetali producono più biofilm e polisaccaridi extracellulari a 28C° rispetto ai ceppi da uomo, oltre che essere differenti per utilizzazione di sorgenti di C, di zuccheri e di sostanze fenoliche e per presenza di catalasi. È possibile che la reiterazione dello stato endofita e la ripetuta esposizione agli stress ambientali in colture industriali su larga scala porti nel medio-lungo periodo alla comparsa di cloni assai adattati ai vegetali e forse meno competitivi con quelli associati all’intestino animale. Questo tipo di effetto è suggerito da esperimenti su topi (8). Delle marcature fenotipiche dei ceppi dei batteri zoopatogeni trovati nei vegetali dovrà tener conto chi si occupa della tracciabilità dei materiali vegetali sospettati di essere vettori dei ceppi isolati dai casi di tossinfezione dell’uomo. Allo stato attuale delle conoscenze sarebbe opportuno ritenere i ceppi riferibili a specie potenzialmente zoopatogene trovate internamente ai vegetali come temporanei, occasionali commensali in attesa di chiarire meglio il loro vero ruolo epidemiologico nelle tossinfezioni.
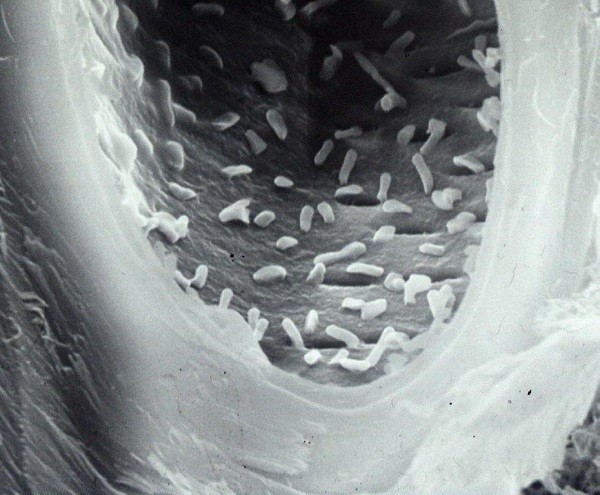
Cellule batteriche endofite all’interno di un vaso xilematico
Umberto Mazzucchi è stato professore ordinario di Patologia Vegetale alla Università degli Studi di Bologna dal 1989 al 2010. Docente del corso di Patologia Vegetale e di alcuni corsi specialistici di materie fitopatologiche, ha svolto ricerche sulle interazioni ospite-patogeno nel processo infettivo e sulla risposta immunitaria delle piante. Pensionato dal 2010, come professore dell’Alma Mater ha svolto seminari occasionali e cicli di lezioni in corsi ufficiali della Università di Bologna.
Lavori citati
1– EFSA, ECDC, Summary Report 2012.
2– Ansa,29 /05/2011; Repubblica,10/06/2011.
3– Guo et al.,2001. Appl. Envir. Microb. 67 (10); Barak et al.,2011. Appl. Envir. Microb 77 (2); Gu et al.,2011. PLoS 6 (11)].
4– Nimchuk et al.,2003. Ann. Rev. Genet. 37.
5– Barak e Schroeder,2012. Ann. Rev. Phytopathol. 50.
6– Seo e Matthews,2012. Appl. Envir. Microb. 78 (16)].
7– Mèric et al.,2012. Envir. Microbiol. 15 (2)].
8– Schikora et al.,2011. PLosOne 6(9).


")
")
")