Note sulla memoria immunitaria delle piante
Prima parte – Memoria immunitaria, un evento epigenetico
di Umberto Mazzucchi

Galla di Ustilago maydis su pannocchia di mais.
Memoria immunitaria, un evento epigenetico
È comune conoscenza che i caratteri ereditari di ogni pianta sono codificati nel suo DNA, lunghissime catene doppie di nucleotidi contenute in cromosomi nel nucleo delle sue cellule. Ogni proteina espressa nella cellula è il risultato di trascrizione e traduzione di una particolare sequenza di nucleotidi (gene) del DNA. La trascrizione ha luogo per mezzo di fattori di trascrizione (proteine) che generano RNA messaggero mobile (catena polinucleotidica; mRNA) che porta l’informazione genetica nei siti delle cellule dove ha luogo la traduzione della sequenza nucleotidica in catene polipeptidiche, basi per la creazione tridimensionale della proteina, espressione terminale della sequenza originale codificata nel DNA. L’insieme delle proteine tradotte ed espresse funzionalmente nei differenti siti cellulari dei vari organi condiziona il fenotipo della pianta.
Per cromatina può definirsi tentativamente nel nucleo di una cellula l’insieme degli acidi nucleici (DNA e RNA) e delle proteine connessi alla trasmissione dei caratteri ereditari codificati dai geni. Della cromatina fanno parte i cromosomi, gruppi di nucleosomi costituiti da pacchetti di 8 proteine (istoni) a due tetrameri sovrapposti (H2A, H2B, H3, H4) attorno cui è avvolto a 2 giri il filamento a catena doppia del DNA (166 paia di basi attorno agli istoni) e i fattori di trascrizione, proteine responsabili della trascrizione della sequenza del DNA di ogni gene nel relativo RNA messaggero (Fig. 1). Un istone H1 è posto ai lati del tratto di DNA di congiungimento tra un nucleosoma e l’altro. I nucleosomi sono strutture dinamiche che rendono il DNA disponibile per la trascrizione. Alterando i nucleosomi è possibile regolare la trascrizione dei geni. Metilazione del DNA e modificazione degli istoni sono gli eventi più comuni messi in atto dalle cellule per regolare la trascrizione; nel primo caso gruppi metilici (CH3-) sono legati a residui di citosina lungo la catena del DNA in modo simmetrico o asimmetrico, nel secondo caso le proteine istoniche vanno soggette sulla coda terminale N a modifiche post- traduzionali con legami di lisina ad acidi grassi a catena breve (es. acilazioni: gruppi R-CO-; acetilazioni: con gruppi acetile, CH3CO-). Queste modificazioni della cromatina sono reversibili, nel senso che ci sono enzimi che legano e altri che distaccano (es. metilasi e de-metilasi per il metile); in particolare, quelle simmetriche del DNA possono essere ereditabili sia attraverso la riproduzione sessuata passando la meiosi, sia con la propagazione vegetativa. Anche RNA non codificanti proteine (ncRNA: siRNA e miRNA) possono contribuire alla metilazione del DNA, a bloccare traduzioni e a variazioni post-trascrizionali e post-traduzionali di proteine. La cromatina può assumere struttura compatta (eterocromatina) o rilassata (eucromatina); nel secondo caso diviene più prona alle trascrizioni e alle tappe successive (2,4,8,10).
Per fenotipo di una pianta in un determinato ambiente si intende l’insieme dei caratteri morfologici e fisiologici espressi da quella pianta in quel particolare ambiente. Ogni carattere espresso deriva dalla traduzione di uno o più geni che le cellule dei vari organi della pianta attuano in quell’ambiente.
Le piante vascolari sono immobili, per lo più fissate nell’ambiente naturale o agrario dove vivono, e sono assai condizionate dai suoi fattori biotici (patogeni, fitofagi, fitofarmaci, inquinanti atmosferici, potature, concimazioni, aggiunta di endofiti radicali, biostimolanti, etc.) e abiotici (siccità, ristagni, alte e basse temperature, intensità luce solare, ventosità, inquinanti atmosferici etc.). Variazioni dei fattori ambientali causano stress alle piante ovvero inducono nelle piante risposte fisiologiche di adattamento alla variazione del fattore(-i) nell’ambiente modificato per poter sopravvivere e possibilmente svolgere il proprio ciclo biologico. Di conseguenza, molteplici sono gli stress che affliggono le piante nel tempo in un dato ambiente. A dire il vero, c’è chi distingue gli stress in positivi o negativi. Così, ad esempio, una appropriata concimazione azotata o l’aggiunta ipogea di rizobatteri simbionti possiamo ammettere causino stress positivi, arrecanti benefici alla pianta. Comunemente, tuttavia, sono gli stress negativi, ovvero “sfavorevoli”, che preoccupano e, infatti, sono questi per lo più oggetto della letteratura specialistica.
Per adattare il proprio fenotipo, la pianta sotto stress deve in qualche modo modificare la espressione del proprio patrimonio genetico. Senza alterare la sequenza DNA dei propri geni, la pianta può perseguire lo scopo adottando un efficace stratagemma: apportare modificazioni della cromatina per alterare tappe della sua espressione in modo da adattare il proprio fenotipo al cambiamento dell’ambiente. La strategia consiste in modifiche del DNA (es. metilazione) e degli istoni (es. acilazioni) associate ad attività di ncRNA. Quando siffatte modificazioni sono ereditabili, divengono oggetto di studio della epigenetica (1). L’applicazione della strategia esprime regolazione epigenetica della espressione genica. Evidenza sperimentale ha messo in luce che in realtà l’espressione del fenotipo di una pianta sia sempre condizionata da regolazione epigenetica della trascrizione connessa sia al mutare dell’ambiente sia allo stato fisiologico della pianta (8). Di conseguenza si comprende come le modificazioni della cromatina a seguito di regolazione epigenetica possano perdurare in modo variabile: ore, giorni o per tutta la vita della pianta fino a passare la meiosi e trasmettersi alla discendenza. La propagazione vegetativa è la via più semplice di conservazione. L’adattamento epigenetico conseguente ad uno stress ha il vantaggio di essere reversibile dato che le modificazioni possono essere cancellate in assenza dello stress.
Una pianta dimostra memoria quando trovandosi sotto stress in ambiente divenuto per qualche aspetto sfavorevole modifica il proprio fenotipo in modo razionale, dettato da esperienza precedente maturata in ambiente analogo, in modo da adattarsi e sopravvivere nel nuovo ambiente. In breve, l’acquisizione della memoria è conseguenza di modificazioni della cromatina avvenute durante la permanenza sotto un certo stress nell’ambiente precedente, conservate nel tempo, ed espresse in occasione del ritorno della pianta sotto lo stesso stress nel nuovo ambiente. Lo stress inducente memoria può essere biotico (es. infezioni di patogeni) o abiotico (es. siccità, alte, basse temperature etc.). Così, ad esempio, l’acquisizione di resistenza di piante alla siccità è conseguenza di memoria generata da stress abiotico: piante che hanno sofferto a lungo per carenza d’acqua divengono più resistenti alla siccità rispetto ad altre che ne hanno sempre avuto normale disponibilità. La memoria per la siccità, marcata da metilazioni del DNA, modifiche di istoni e alterazioni post-trascrizionali e post-traduzionali, perdura poco e non si trasmette alla progenie (3). Questo tipo di memoria, definita epigenetica, è stata evidenziata anche nelle cellule animali (5).
La regolazione epigenetica della espressione genica è molto diffusa sia negli eucarioti che nei procarioti, tenuto conto della diversità di struttura dei loro genomi. Nei batteri, ad esempio, si ha metilazione della adenina o della citosina e, in assenza di istoni, le modifiche hanno luogo per mezzo di proteine particolari NAPs associate al DNA (6)
I progressi della epigenetica nell’ultimo decennio stanno offrendo nuovi spunti per il miglioramento genetico delle piante agrarie (3,7). Superare o contrastare con successo le difficoltà dell’ambiente in cui vive in modo razionale è anche manifestazione di intelligenza delle piante!
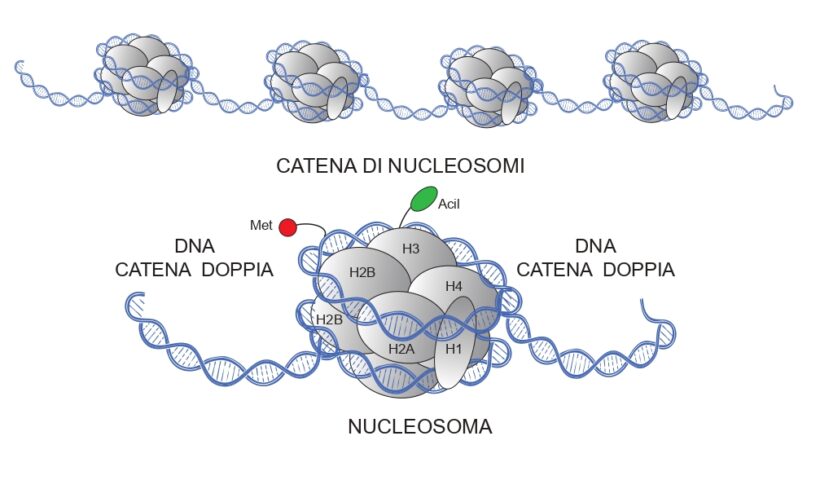
Fig.1. Schema della struttura di un nucleosoma e della loro disposizione lungo la catena doppia di DNA. Ogni nucleosoma consta di un ottamero di istoni (proteine) (H2A, H2B, H3, H4)2 più un istone H1 stabilizzatore, e ha attorno due giri di DNA a catena doppia (166 paia di basi). Le catene di nucleosomi si aggregano poi in solenoidi, stabilizzati dalle H1, che si comprimono ulteriormente a formare i cromosomi. Metilazione del DNA ( Met ) e acilazione (Acil ) di un istone sono abbozzati.
Lavori citati
1- Deichmann U.,2016. Development Biology 416,1,249-254.
2-Nakamura K. et al.,2025. Current Opinion in Plant Biology 87,102763,1-9.
3- Kambona C.M. et al.,2023. Theoretical and Applied Genetics 136,26,1-21.
4- Kashyap S. et al.,2024. Plant Science 340,111954,1-11.
5- Palacios S. et al.,2025. Cell Genomics 5,100985,1-14.
6- Verma S.C. et al.,2019 Plos Genetics 12, e1008456,1-35.
7- Wang X. e N. Yamaguchi, 2024.Current Opinion in Plant Biology 81,102559,1-9.
8- Xie S-S. e C-G. Duan, 2023.aBiotech 4,124-139.
9- Yamamuro C. et al., 2016.Molecular Plant 9,57-76.
10- Zang X., 2008.Science 320,489-492.
Glossario
ncRNA : Acido ribonucleico non codificante.
mRNA : Acido ribonucleico messaggero.
siRNA : Acido ribonucleico di interferenza.
PTI: Pattern Triggered Immunity- Prima fase della risposta immunitaria innata delle piante.
ETI: Effector Triggered Immunity- Seconda fase della risposta immunitaria innata delle piante (integra PTI).
Non-self: Una biomolecola o un organismo che la pianta percepisce essere di diversa origine o in condizione
differente dal normale, riconosciuti durante la propria attività metabolica per conservare la
omeostasi.
Riconoscimento in negativo: Una cellula tende ad accettare e a integrarsi funzionalmente con l’entità
non-self fino a che non riceve un segnale di incompatibilità.
Riconoscimento in positivo: Una cellula continua a considerare la entità come non-self e non si integra
fino a che non riceve un segnale positivo di compatibilità.
Umberto Mazzucchi è stato professore ordinario di Patologia Vegetale alla Università degli Studi di Bologna dal 1989 al 2010. Docente del corso di Patologia Vegetale e di alcuni corsi specialistici di materie fitopatologiche, ha svolto ricerche sulle interazioni ospite-patogeno nel processo infettivo e sulla risposta immunitaria delle piante.
Pensionato dal 2010, come professore dell’Alma Mater ha svolto seminari occasionali e cicli di lezioni in corsi ufficiali della Università di Bologna.