Storia Naturale della vita sul pianeta Terra
Tentativo di una sintesi
di Giuliano Russini
Dicono che il mare sia freddo, ma il mare ha il sangue più caldo che esista – da: Le balene non piangono – D.H. Lawrence, 1885-1930
In questo articolo, vorrei provare a tracciare per grandi linee, riassumendo diversi concetti tassonomici, evolutivi, paleontologici e biogeografici, come la vita, una volta nata, si è evoluta e diversificata sulla Terra.
Lungi da me voler fare un’opera esaustiva, anche solo pensarlo, sia perché lo spazio a disposizione non ce lo consente (occorrerebbero decine di volumi, ciascuno di un migliaio di pagine, per avere un quadro appena generale), ma anche perché pur essendo un biologo zoologo-etnogeografo, come formazione, gli aspetti evolutivi e di Storia Naturale per tutti gli organismi ripartiti nei cinque Regni (animale, vegetale, funghi, protisti e monera o procarioti, a cui si aggiungerebbe un sesto non biologico, il Regno minerale), non sono per me allo stesso livello di conoscenza; ad esempio sono obiettivamente più carente nei Procarioti (Procariota) e infine, cosa più importante, ci sono biologi molto prestigiosi e accademicamente qualificati a livello internazionale e nazionale che hanno sicuramente la cultura per poterlo fare molto meglio.
Non verranno considerate in questo articolo neanche tutte quelle teorie più o meno scientifiche, in alcuni casi fantasiose, che tentano di tracciare la nascita della vita sul pianeta, quindi su come sia nata la vita, non come poi si sia evoluta darwinianamente; una su tutte la teoria del cosiddetto “Mondo a RNA” che, come altre ancora, non ricadono nella biologia sistemica ma hanno più una matrice chimico-fisica, totalmente al di fuori delle mie competenze.
Lo scopo di questo articolo, invece, è mostrare come il biologo, sia esso uno zoologo, un botanico, un ecologo, un etnobiologo-antropologo, un paleontologo o un paleobiologo, imposti il suo ragionamento sulla Storia Naturale di una specie e su come la vita si sia struttura e si sia diversificata (e continui a farlo) in tutte le sue eteroforme nel corso dei tempi geologici; questo perché, se il fisico è il più intimo conoscitore delle leggi elementari della natura (dove per elementare s’intende generale, come la forza di gravità), il chimico della materia, il biologo è il più profondo conoscitore della Storia Naturale della vita sul pianeta Terra e dell’avventuroso viaggio che essa ha affrontato e affronta.
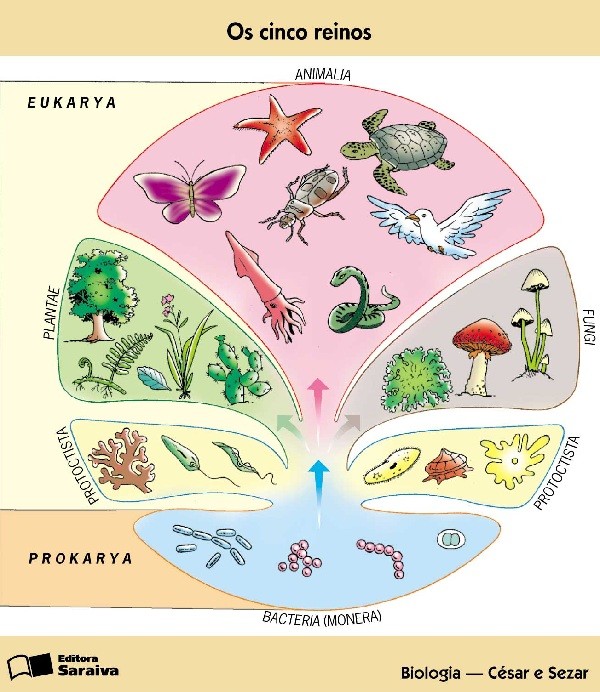
Foto 1. – I cinque Regni, da Biologia di César-Sezar
Il Quadro generale
Possiamo iniziare, considerando le massime suddivisioni nel mondo vivente, che sono chiamate
“Regni”.
Tutti conoscono il Regno animale e il Regno vegetale.
In senso lato, essi vengono principalmente distinti dalla fondamentale differenza che esiste tra
l’autotrofia dei vegetali, consentita dalla chemiosintesi e soprattutto dalla fotosintesi e, l’eterotrofia
degli animali che, non potendo effettuare una chemiosintesi, dipendono in gran parte per il loro
sostentamento dalla fotosintesi dei vegetali.
Questa suddivisione nei due regni non è del tutto soddisfacente per il biologo.
Per esempio, i funghi sono tutti eterotrofi e la loro struttura è sinciziale, hanno cioè molti nuclei non divisi da pareti e membrane cellulari: non sono quindi a ragione oggi, a differenza di quanto avveniva in passato, posti nel Regno Vegetale (Plantae), ma ne hanno un proprio, ovvero il Regno dei Funghi (Fungi).
Oppure, fino agli anni ’40 del secolo scorso, alcuni li collocavano anche nel generico Regno degli Eucarioti (Eukaryota).
Il gruppo molto omogeneo dei Flagellati (Flagellata) comprende forme autotrofe con clorofilla e forme eterotrofe che ne sono sprovviste, anche nell’ambito della stessa famiglia.
I biologi quindi sono stati indotti a rivedere quella che è stata la definizione di Regno e a inserirvi anche delle considerazioni strutturali.
Aggiungeremo quindi alla definizione del Regno vegetale e del Regno animale, che si tratta di eucarioti, cioè di forme con nucleo individualizzato.
Una tale aggiunta permise di distinguere e creare un altro Regno, quello dei Procarioti o Monere (Procariota, detto anche delle Monera) o a nucleo diffuso, cioè delle Schizofite (Schizophyta), che non hanno raggiunto la capacità di riprodursi sessualmente.
Un’ulteriore precisazione alla definizione di Regno consiste nel fatto che i Regni vegetale e animale non sono ben distinti, tranne che nelle forme pluricellulari, il che individualizza il Regno dei Protisti (Protista), comprendente tutti gli eucarioti monocellulari, autotrofi e no.
E’ ampiamente giustificato raggruppare i protisti in un Regno a parte.
Essi possiedono infatti organuli, o organiti e hanno non solo una bocca e dei vacuoli escretori, come nei ciliati, ma in alcuni flagellati (Warnowidi) l’ultramicroscopia ha permesso di riconoscere strutture oculari complesse con cornea, cristallino e retina, a cui vanno aggiunti organi urticanti per la difesa, paragonabili alle cnidocisti dei celenterati.
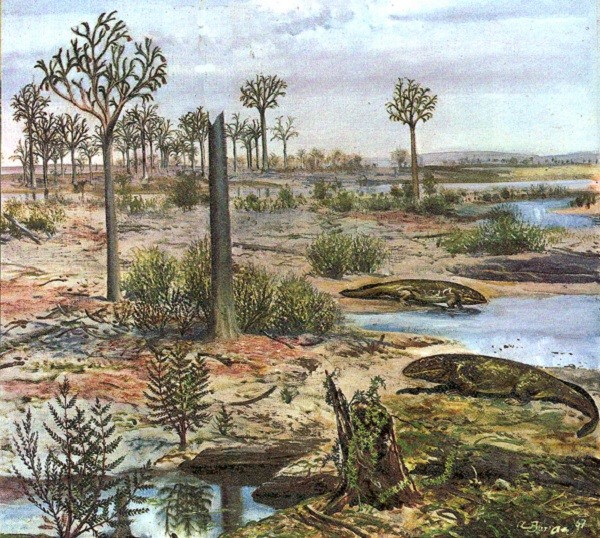
Foto 2. – Paleoambiente del Devoniano, conquista dell’ambiente terrestre da parte delle piante, da “Atlas de Biologie préhistorique et Paléontologie”
Esaminiamo ora, la Storia Naturale dei singoli Regni.
Biocinesi del Regno dei Procarioti (schizofite)
A partire dal supereone Precambriano o Archeozoico (il supereone, è l’unità di tempo geologico più ampia), più di quattro miliardi e mezzo di anni fa, i procarioti, che corrispondono forse alla “monere” del biologo Ernest Heckel (1866), sembrano essersi evoluti molto poco nella morfologia.
Le Cianoschizofite (Cyanoschizophyta) hanno avuto le stesse forme e sono giunte alle medesime concrezioni calcaree di oggi.
Sembra anche che le Batterioschizofite (Bacterioschizophyta) non abbiano subito alcuna evoluzione morfologica, dopo il Precambriano, anche se la loro funzionalità biochimica ha potuto evolversi.
Inoltre, è possibile che in alcune condizioni i batteri della stessa specie siano in grado si effettuare la sintesi di diverse sostanze.
Biocinesi del Regno dei Protisti
I flagellati, nel loro insieme, effettuano sintesi biochimiche capaci di caratterizzare ogni famiglia.
Alcuni, provvisti di uno scheletro, sono morfologicamente ben definiti.
I Rizopodi (Rhizopoda) e i Foraminiferi (Foraminifera) hanno caratteristiche che permettono ai biologi di ricostruirne la Storia Naturale, che tuttavia viene resa più complessa dalla presenza, nei cicli riproduttivi, di fasi sessuate (con dimorfismo), produttrici di gameti e di fase asessuate, generatrici di spore, cicli “metagenetici alternati”.
Inoltre, la morfologia è molto sensibile alle variazioni di temperatura, di salinità, di profondità, tanto che le differenze che potrebbero essere attribuite a specie e anche generi distinti, sono solamente una sorta di “sommazione somatica” o “somazione”: come accade nei Coccolitoforidi (Coccolitoforidae) e nei Foraminiferi (Foraminifera).
Biocinesi del Regno vegetale
Se si escludono i protocarioti e i protisti, il Regno vegetale comprende una serie di gruppi algali (che risalirebbero da resti fossiliferi, a circa 1,6 miliardi di anni fa!) che si distinguono a prima vista per i pigmenti che possono accompagnarsi alla clorofilla: pigmenti bruni delle Feoficee (Phaeophyceae), pigmenti rossi delle Rodoficee (Rhodophyceae), mentre nelle Cloroficee (Chlorophyiceae) la clorofilla è presente da sola.
E’ indubbio, quindi, che la strada venne aperta da organismi vegetali unicellulari e, che come è ben noto, la vita nacque grazie all’acqua e “nell’acqua”.
Questi tre gruppi algali, differiscono per il metabolismo e possono essere considerati affini ai protisti flagellati, già differenziati gli uni dagli altri.
Le feoficee (alghe brune) e le rodoficee (alghe rosse) si sono evolute separatamente fino a divenire forme molto avanzate, pur rimanendo sempre alghe, cioè sempre sprovviste di sistemi vascolari e sempre in stretto rapporto con l’ambiente acquatico (marino e dulcacquicolo); la vita in acqua, necessaria alla riproduzione, garantisce anche un apporto di nutrienti che svincola dalla necessità anatomica di un apparato vascolare, per cui sono tutti organismi avascolari.
Occorre osservare tuttavia, che alcune alghe brune sono lunghe anche decine di metri e che hanno
saputo adattarsi isolatamente alla zona “tidale”, la piana di marea, o “sopratidale”, la zona al di sopra del livello medio di alta marea, acquisendo in maniera del tutto occasionale, una sorta di protovascolarizzazione.
E’ questo il caso delle Prototaxites (del periodo Devoniano inferiore, era Primaria o Paleozoica, della penisola Gaspé o Gaspesie, sudest del Quebec, Canada) e di Crocalophyton (periodo Devoniano superiore, era Paleozoica, dell’America settentrionale).
Le cloroficee, hanno subito una evoluzione completamente diversa e è interessante sottolineare che solo un phylum di questo antico gruppo di alghe si è realmente adattato alla vita subaerea.
Questo adattamento, si riscontra nelle Tracheofite (Tracheophyta), al cui gruppo appartengono tutti i vegetali terrestri.
La comparsa e l’evoluzione delle piante tracheofite pose e pone numerosi problemi ai biologi.
Siamo nel periodo Devoniano, circa 417 milioni di anni fa, uno dei periodi in cui è suddivisa l’era Paleozoica o Primaria, che a sua volta afferisce all’eone Fanerozoico, così denominato per la comparsa di forme viventi pluricellulari, in contrapposizione al supereone Archeozoico o Precambriano, ove si ebbe la comparsa di prime protoforme di vita marina, per lo più spongiari, protozoi ecc.; il nome di Devoniano venne assegnato dai geologi a questo periodo, usando il nome della contea inglese di Devon, ove gli affioramenti di rocce di questo periodo sono abbondanti.
In questo periodo, comincia ad aversi il distacco sempre più progressivo delle forme vegetali, a cui fanno parallelamente supporto modificazioni anatomiche, che via, via, generano gruppi di piante sempre più adatte alla vita terrestre o subaerea.
La comparsa di tessuti differenziati si è realizzata progressivamente.
I tessuti conduttori (legno e libro o xilema e floema), sono comparsi nelle prime licopodine del Devoniano inferiore (416-398 milioni di anni fa), famiglia Lycopodiales e, contemporaneamente, anche nelle Psilofite (Psilophyta), una divisione di piante primitive, comprendenti le forme vascolari più elementari. Le radici non sono differenziate dai gambi sotterranei, con peluzzi assorbenti. Nelle Psilofite non esistono foglie vere e proprie.
Gli apparati riproduttori (sporangi) sono già localizzati su organi specializzati.
Successivamente, in specie afferenti a varie famiglie come i licopodi, i lepidodendri (che nel Devoniano si presentavano con alberi altri 30 m, con tronchi larghi anche 2 m), le selaginelle e gli equiseti, nascono nuove e più perfezionate strutture anatomiche.
L’apparato fogliare si è già diversificato e un notevole aumento del numero delle foglie, incrementa la superficie esposta alla luce e quindi alla fotosintesi.
Il massimo sviluppo delle foglie a fronde è raggiunto nelle Pteridofite (Pteridophyta) e nelle Pteridosperme (Pteridospermae), a cui appartengono le Felci, classe Filices in latu sensu, ovvero forme di piante preistoriche che si avvicinavano molto alle felci attuali, di cui abbiamo buoni resti fossiliferi, non così comuni per le piante.
Le felci oggi sono nel loro massimo sviluppo evolutivo, si pensi alle forme arboree presenti nelle foreste pluviali della fascia tropicale e subtropicale.
La tappa seguente, o concomitante, è stato il perfezionamento della riproduzione che passa a partire alcune pteridofite, da una fase a spore uguali (isospora), a una fase a spore diverse (eterospora), a cui sebbene raramente e in pochi gruppi, incomincia ad afferire, transitoriamente, uno stato apogamico (sessi separati); permane invece una condizione aplostelica (un unico stele), per la struttura del caule, nella fase giovanile dello sviluppo; mentre in altri gruppi, tale condizione è più complessa, per cui la struttura del caule, può essere “sifonostelica” o “aplomeristelica”, comunque per la prima volta, nelle piante, si ha un membro tricormo, cioè in cui si distinguono le radici dal fusto e infine questo dalle foglie; tale modificazione, fu una delle chiavi di volta per la conquista dell’ambiente terrestre, in maniera permanente.
Le pteridosperme, come le più antiche pteridofite, possono riprodursi anche asessualmente, mediante “propaguli” vegetativi.
Nel passo successivo (parliamo sempre di tempi geologici), le microspore divengono quindi “polline” e le macrospore “ovuli”.
Il passaggio, da una riproduzione a spore ad una mediante gameti di sessi opposti, nel Regno vegetale, è paragonabile nella sua importanza evolutiva, al passaggio dalla oviparità dei pesci o degli anfibi, ove si necessita sempre dell’acqua, alla oviparità mediante “uovo cleidoico” nei rettili, per il Regno animale; ciò che permise in via definitiva agli animali, di abbandonare in maniera permanente l’ambiente acquatico e svolgere il loro ciclo vitale sulla terraferma.
Tale conquista, conclamatasi pienamente nelle pteridosperme durante il Devoniano, le affranca quindi definitivamente dall’ambiente acquatico, sia rispetto le pteridofite ove tale processo era solo nella fase iniziale, infatti questo gruppo di piante dipendeva ancora dall’acqua per la riproduzione mediante gameti flagellati, sia rispetto le preistoriche Briofite (Bryophyta) come muschi, epatiche e antocerote e ancor più rispetto le alghe, che dipendevano e dipendono totalmente dall’acqua.
Passiamo quindi da piante in cui domina il “tallo” o lamina, perciò dette tallofite, che sono totalmente acquatiche, a piante in cui domina il “cormo” ovvero fusto, tipico delle piante terrestri.
Con le pteridosperme, si esce quindi dal campo delle crittogame vascolari e si può cominciare a parlare di “prefanerogame” o “protofanerogame”.
L’ovulo fecondato via, via, acquisisce sostanze di riserva e diviene un seme, affrancato dalle avverse condizioni ambientali e climatiche e dalle temporanee siccità.
E’ lo stadio delle Fanerogame (Fanerogama), o Spermatofite (Spermatophyta), il biologo Linneo le chiamava Antofite (Anthophyta,cioè pianta con fiore); le Spermatofite (Spermatophyta), sono un ordine evolutivamente moderno, o più in alto nella scala botanica, rispetto il superordine delle Archegoniate (Archegonyatae), formato dalle Briofite (Bryophyta) e le Crittogame (Cryptogame) come le Pteridosperme (Pteridospermae); le spermatofite ovvero piante a seme, hanno nelle Gimnosperme (Gymnosperma), cioè piante con ovuli nudi, le prime piante afferenti a questo gruppo, comparse sul pianeta, mentre solo in seguito sono comparse le Angiosperme (Angyosperma), piante con ovuli rivestiti, in cui evolutivamente compare anche il fiore perfetto; faccio notare che, alcuni biologi ancora oggi, considerano fiore anche quello delle gimnosperme, ad esempio lo “strobilo” delle conifere o delle Cicadili (Cycadeles), mentre altri autori solo quello delle angiosperme.
Il fiore della Gimnosperme, è privo di perianzio e delle relative aree o camere di accettazione-relazione e distribuzione per gli insetti pronubi, quindi l’impollinazione è assente, in particolare l’entomofila.
Il fiore delle Angiosperme (quello di Magnolia è considerato il fiore primitivo) si dota di perianzio (ovvero calice più corona), di petali e sepali colorati, ove sono tracciate in molte specie le linee del miele, tutte caratteristiche che sono coevolute con la classe degli Insetti (Insecta), per la formazione degli Imenotteri (Hymenoptera), Lepidotteri (Lepidoptera) come anche diversi Coleotteri (Coleoptera), pronubi, necessari alla riproduzione mediante impollinazione; tale fiore può essere diclino o dioico cioè o maschile o femminile (quindi dotato o solo di stami, organo riproduttivo maschile, o solo di pistilli, l’organo riproduttivo femminile), o monoclino o monoico, ovvero ermafrodita, cioè quel fiore nel quale sono presenti gli organi sessuali di entrambi i sessi; da cui una pianta si dice monoica, quando produce i fiori di entrambi i sessi, dioica quando produce fiori o solo maschili o solo femminili.
La riproduzione in queste piante, avviene quasi sempre mediante il fiore, raramente per propagazione, più tipica nelle Gimnosperme.
Un’altra strategia di riproduzione per molte angiosperme si conclama con la produzione di diversi tipi di frutti partendo dal fiore, che ne rappresenta la fase vegetativa, i quali mangiati da animali frugivori, ne garantisce la propagazione dei semi mediante le feci.
I vegetali a foglie caduche (che cadono), come le latifoglie per gli alberi, in grado di resistere particolarmente bene all’alternarsi delle stagioni, grazie a periodi di pausa invernale, appaiono all’apice della biocinesi del Regno vegetale.
Si noti il carattere stratificato di questa biocinesi delle clorofite, che indica una graduale conquista dell’ambiente subaereo:
- Accelerazione della fotosintesi, grazie allo sviluppo fogliare.
- Isolamento in seno all’ambiente esterno, grazie alla comparsa dapprima di semi sempre meglio protetti e in seguito, dalla pausa invernale dei vegetali a foglie caduche.
E’ probabile che ogni tappa sia stata raggiunta nelle diverse linee evolutive in seguito a fenomeni d’evoluzione parallela. Ognuna di queste tappe comprende al suo interno una serie di forme che hanno raggiunto lo stesso grado di complessità. Si tratta quindi, in questo caso, di una evoluzione seriale.
Biocinesi del Regno Animale
Il Regno animale, eccezion fatta per i protisti di tipo animale, ha subito una biocinesi (evoluzione), che è egualmente complessa o forse anche più a quella vegetale, poiché si è basata molto anche sul differenziamento dei tessuti.
Infatti, mentre nelle tracheofite l’insieme dei tessuti permane a livello vascolare, nel Regno animale il differenziamento degli organi si inserisce in un panorama molto più vasto e vario, che comporta elementi molto diversi.
Si possono distinguere, le stratificazioni o tappe dell’evoluzione animale, dapprima in rapporto al numero e alla posizione degli strati cellulari, o foglietti, dai quali traggono origine gli abbozzi embrionali.
Il loro numero varia da uno a tre.
1. Nella fase dei “monoblastici”, l’animale possiede solo un tipo di tessuto, un solo foglietto embrionale.
Questo stadio, rimane ipotetico per quanto riguarda gli animali attuali, ma sembra essere stato bene individualizzato all’inizio delle ere paleontologiche, per esempio con gli Archeociati (Archaeocyatha), metazoi primitivi esclusivi dei mari del Cambriano e molti altri gruppi, oggi completamente estinti.
2. Nella fase dei “diblastici” (o diploblastici di Ray Lankaster, 1873, il biologo che li studiò), si sono differenziati due foglietti: l’ectoderma e l’endoderma.
Restano talvolta nella fase di strati in cui le cellule mantengono una certa indipendenza.
E’ il caso delle spugne, nelle quali non è possibile distinguere tessuti veri e propri.
Al contrario, nei celenterati l’ectoderma e l’endoderma, sono ben differenziati a formare dei tessuti e racchiudono anche sistemi sensitivi e motori.
In ogni caso l’ectoderma presenta cellule differenziate nel senso della sensibilità, della protezione e della difesa; l’endoderma, presenta cellule differenziate nel senso dell’assorbimento e dell’assimilazione.
Tra i due, s’inserisce uno strato gelatinoso (la mesoglea) in sé anisto (cioè privo di una struttura determinata), ma che racchiude gli elementi di un cinesoma, cioè di uno scheletro in forma di armatura e di fibre contrattili.
Il cinesoma muscolare è ben sviluppato nelle meduse.
In questa fase, si riscontrano ancora caratteri indifferenziati, per esempio la rigenerazione a partire da frammenti sezionati.
Inoltre i celenterati più primitivi Idrozoi (Hydrozoa), presentano alternanza di generazioni: polipi fissi e meduse libere.
Queste due forme, coincidono anche con le due estremità tra le quali si è sviluppata l’evoluzione seriale del gruppo: polo-medusa, scifozoi; polo-polipo, antozoi.
Questa situazione deriva indubbiamente da un’antica evoluzione parallela.
Il differenziamento di organi per una funzione determinata, ha preso negli idrozoi l’aspetto di un differenziamento di vari individui raggruppati in una colonia.
Effettivamente le varie funzioni come nutrimento, difesa e riproduzione, sono ripartite fra tre forme di polipo differenti, che coesistono in un numero indeterminato di esemplari in uno stesso insieme, dove si trovano legati l’uno all’altro da una base comune, il “cenosarco”.
In altri Celenterati (Coelenterata), un polipo o una medusa, possono individualmente assumersi le diverse funzioni.
3- La fase dei “triblastici acelomati” (il termine triploblastici fu coniato nel 1873 da Ray Lankester,
preferisco usare quello più eufonico per la nostra bella lingua, di origine francese di “triblastico”), è caratterizzata dalla presenza di un terzo foglietto embrionale, il mesoderma, che normalmente deriva dall’endoderma, ma è inserito tra questo e l’ectoderma.
Un abbozzo del mesoderma, si riscontra nei gruppi più diversi che, secondo i casi, si possono avvicinare ai diblastici, come gli Ctenofori (Ctenophora) o gli Anellidi (Anellida), ad esempio i Turbellari (Turbellaria).
A questo livello, si colloca un certo numero di forme intermedie, di difficile classificazione per i biologi tassonomisti -in particolare, la maggior parte dei vermi parassiti per l’essere umano, gli animali, come i Platelminti (Platyhelminthes) e Nematelminti (Nemthelminthes) – nelle quali si osserva una prima tendenza alla ripetizione metamerica degli organi.
4. I “triblastici celomati” possono essere considerati come uno stadio più avanzato: in essi il mesoderma, si sviluppa e si scava, originando una cavità interna molto importante, il celoma o cavità generale. Il celoma, pieno di un liquido che non può uscirne, ha un compito di primo piano nel cinesoma, come scheletro idrostatico.
Ma ospita anche i diversi organi, in particolare quelli escretori, i “nefridi” e le gonadi.
Quanto al mesoderma, esso ha ereditato la maggior parte delle prerogative proprie alla mesoglea dei diblastici, racchiude, cioè, anche elementi del cinesoma: fibre muscolari e scheletro assiale.
Questa fase dello sviluppo evolutivo e della Storia Naturale dei celomati, comprende gli elementi principali della fauna attuale fossile. Tali elementi possono essere riuniti in serie divergenti a partire da un punto centrale che, grosso modo, corrisponde agli anellidi.
In questa serie, non esistono passaggi laterali possibili, ma solo evoluzioni parallele a partire da forme che dovevano essere molto affini alla larva “trocofora” degli anellidi.
Sembra che l’ontogenesi (l’evoluzione dell’embrione), sia stata responsabile, nel caso in cui le fasi adulte sono comparabili (per cui sono dette linfoidi) della maggior parte delle strutture che distinguono fra loro i gruppi animali principali, alcune rimanendo a tre segmenti larvali (trimetameri) e le altre moltiplicando il numero dei segmenti nella larva compiuta o nell’adulto, come nel caso degli anellidi, dei molluschi arcaici e degli artropodi.
5. In tre Phyla, un nuovo progresso giustifica la distinzione di uno stadio dei “cefalizzati”.
Sono quelli la cui porzione anteriore, che porta non solo la bocca, ma anche diversi organi di senso, si sviluppa e coinvolge la maggior parte dei segmenti anteriori che si fondono più o meno completamente.
Questi tre Phyla, sono quelli degli Atropodi (Arthropoda), Molluschi (Mollusca) e dei Cordati (Chordata).
Alcune tendenze di cefalizzazione si trovano negli anellidi e forse altrove, per esempio negli Echinodermi (Echinodermata), riferiti ai Calcicordati (Calcichordata) dal biologo Jeffries nel 1937.
Dai cordati in poi si forma quella lunga serie ramificata di Vertebrati (Vertebrata) che, tra il periodo Carbonifero superiore o Pennsylvaniano e il periodo Permiano dell’era Paleozoica o Primaria e il periodo Trias o Triassico dell’era Mesozoica, portò alla nascita dei rettili, i quali sebbene non furono i primi vertebrati tetrapodi terrestri, poiché pur se in comunione ancora con l’ambiente acquatico per la riproduzione, i primi vertebrati tetrapodi terrestri furono gli Anfibi (Amphibia), a loro volta derivati da un sottordine estinto di pesci i Rhipidistia, furono però quelli che per primi divennero totalmente indipendenti dall’acqua, sin dalle prime forme di Cotilosauri (Cotylosauria), grazie al già citato uovo cleidoico.
Da qui si ebbe nell’evoluzione dei rettili, la nascita dei Dinosauri, il cui massimo splendore coincide con il Mesozoico intermedio, periodo Giurese o Giurassico.
Dopo la loro estinzione, l’evoluzione dei rettili continuò con la nascita di altri gruppi, ma intanto piccoli animali endotermi-stenotermi, dotati della capacità di allattare mediante mammelle i propri nascituri, cioè i Mammiferi (Mammalia), dapprima rappresentati da animali grandi quanto un topo, si andavano sempre più affermando, conquistando tutte le nicchie ecologiche via, via disponibili, modificandosi e avendo la massima variazione evolutiva durante il Cenozoico o era Terziaria.
Senza scordarci né l’evoluzione della classe degli Insetti (Insecta), come di tutti gli artropodi, né del gruppo dei Pesci (Pisces), o della importantissima classe degli Uccelli (Aves), che deriva probabilmente dai rettili, in stretto legame con dinosauri, al punto che alcuni biologi paleontologi definiscono gli uccelli i “dinosauri” moderni.

Passaggio da un Regno all’altro
Tutto quanto è noto sull’origine del mondo vivente fornisce prove che permettono di presumere che il differenziamento dei Regni si sia realizzato nel supereone Precambriano.
Le tappe intermedie, che ci permetterebbero d’acquisire alcune idee sul loro modo di realizzarsi, sono dunque molto rare e di dubbia interpretazione.
Il passaggio dall’autotrofia clorofilliana all’eterotrofia animale si è accompagnato allo sviluppo della facoltà del movimento, che è tuttavia presente anche nel Regno vegetale, per esempio nel caso dei gameti flagellati.
L’esistenza di colonie di protisti, quali i dinoflagellati policrichidi e Haplozoon o quali i volvocidi, ha da tempo mostrato come si sono originati i Metazoi (Metazoa).
Rizopodi, quali i foraminiferi, presentano stadi sinciziali e pluricellulari, dimostrando caratteri che potrebbero farli migrare da un Regno all’altro.
Conclusioni
Concludo questa breve e maldestra sintesi della Storia Naturale della vita sul nostro pianeta, partendo dalla frase citata nell’introduzione, di D.H. Lawrence, grande e virtuoso scrittore britannico: “Dicono che il mare sia freddo, ma il mare ha il sangue più caldo che esista”, per ricordare a tutti noi, che se siamo vivi oggi e lo saremo ancora per altre generazioni, se questo Pianeta è potuto essere un’opera di bellezza, questo è stato grazie all’acqua, che sin da quando siamo feti, in un sacco amniotico, ci accompagna e poi ci accompagnerà per tutta la nostra vita; l’acqua che è la nostra più dolce sorella, verso cui purtroppo non abbiamo più alcun rispetto, né considerazione, come d’altronde verso la biosfera in cui viviamo.
La vita (e la sua Storia Naturale), questa meravigliosa contraddizione pulsante più di qualsiasi fenomeno nell’Universo, rappresenta per il biologo un labirinto entro cui si perde continuamente, per la sua inafferrabile e mutevole complessità, ma in questo caso non c’è nessun filo d’Arianna a guidare questo Teseo verso la luce.
Giuliano Russini è laureato in Scienze Biologiche all’Università La Sapienza di Roma, con specializzazione in botanica e zoologia; successivamente ha conseguito in UK e Francia la specializzazione in etnobiogeografia. Lavora come curatore al Giardino Esotico di Hendaye, Francia. (e-mail: russinigiuliano@yahoo.it).
 |
Ebook – La Prossima Era Glaciale Scopri cos’ha provocato l’estinzione dei dinosauri e perchè potrebbe presto minacciare la nostra sopravvivenza… |